Fig. 2
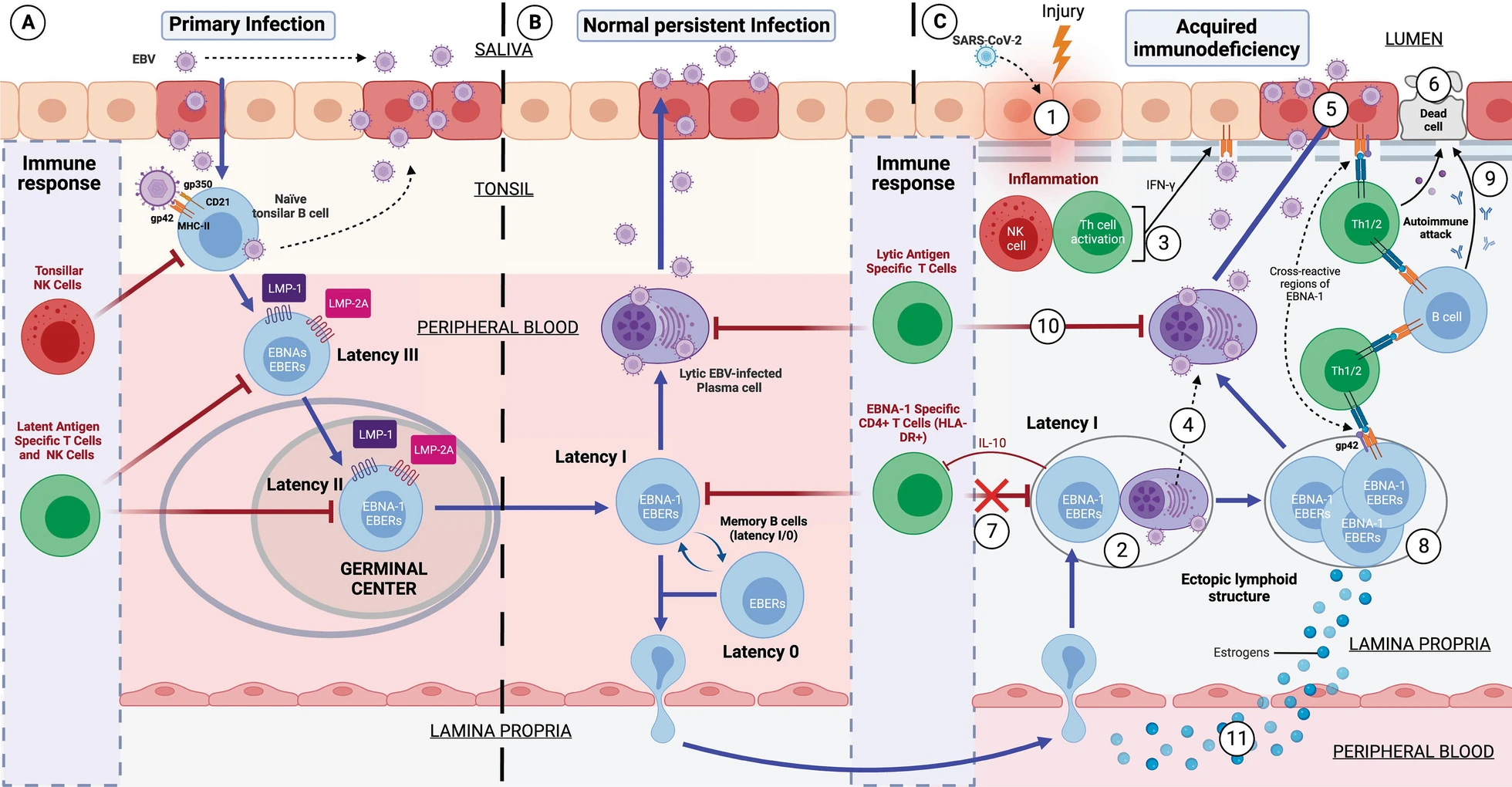
Development of Epstein–Barr Virus (EBV)-induced acquired immunodeficiency in patients with genetic susceptibility. A Primary infection by EBV. EBV is transferred to the host via the saliva of an infected carrier, initially infecting the epithelial cells of the pharynx and subsequently naïve B lymphocytes in the tonsils, through interactions between the virus’s glycoproteins gp350 and gp42 and the host cells’ CD21 and Class II MHC (MHC-II) molecules, respectively. Lytic infection creates new viral particles that continue to infect more epithelial cells. Subsequently, the EBV-infected B cells enter a state of peripheral latency, during which they express a set of specific viral genes, including LMP1, LMP2A, EBNAs, and EBER (latency III). These latency III B cells progress through the germinal center reaction to latency II and emerge as memory B cells with latencies I/0 that establish a lifelong latent infection (B). The healthy host’s immune response is sufficient to control the EBV infection. NK cells in tonsil produce high levels of IFN-γ that withhold the transformation of B cells by EBV during earlier stages of the infection. Type III and II latency B cells are regulated by NK and T cells specific for latent proteins. However, memory B cells with type I latency are only controlled by activated EBNA-1 specific CD4 T cells. EBV-infected plasma cells can periodically enter in lytic phase, but are controlled by CD4 and CD8 T cells with specificity for EBV lytic proteins. C Development of EBV-induced acquired immunodeficiency and autoimmunity in the mucosa of genetically susceptible patients. (1) An inflammatory stimulus or other infection (SARS-CoV-2) recruit leukocytes in the mucosa, including latency I (EBNA-1) B cells and healthy B cells. (2) In the mucosa, B cells form ectopic lymphoid aggregates that enable the generation of antigen-specific immune responses. These ectopic lymphoid structures create a favorable environment for the transformation of EBV latent B cells into proliferating blasts, to become memory B cells. (3) Furthermore, activation of NK cells occurs in response to both the initial inflammatory stimulus and to restrict B cell transformation by EBV. Exposure to foreign antigens from the initial stimulus or to viral antigens from EBV leads to the activation of CD4 T cells and the release of IFN-γ, followed by upregulation of MHC-II on epithelial cells, promoting the acquisition of a non-professional antigen-presenting cell phenotype. (4) Additionally, the presence of foreign antigens could also trigger terminal differentiation and activation of latent EBV B lymphocytes, allowing the transition from the latent phase to the lytic virus phase. (5) Subsequently, newly generated viral particles infect more epithelial cells through gp42/MHC-II interaction, leading to increased inflammation and ultimately to latent EBV infection. Moreover, this chronic inflammation induces a cytokine response, leading to further recruitment of B cells and perpetuation of viral infection. (6) Latent EBV epithelial cells could enter a lytic phase, releasing new virions, undergoing lysis due to T cell response, or experiencing neoplastic transformation. (7) Immune evasion mechanisms of EBV latency (epithelial and B cells) involve a decrease in activation and cytotoxic capability of EBNA-1-specific CD4 T cells through the release of IL-10 and EBV miRNAs contained in exosomes, which could suppress the expression of target genes in the viral or host genome to maintain latent EBV infection. (8) This altered immunosurveillance leads to increased proliferation of EBV-latent B- and epithelial cells, which raises the risk of neoplastic transformation or autoimmune disease in genetically predisposed patients with ancestral MHC-II alleles susceptible to EBV. (9) The presentation of native cellular autoantigens or viral EBNA-1 through MHC-II/gp42, which can undergo post-translational modifications, such as citrullination, and form neoantigens, could trigger the activation of autoreactive CD4 T cells and the formation of autoantibodies against tissue cells. (10) Other virus latency phases or the lytic phase would be controlled by NK and CD4 and CD8 T cells, specific for the EBV lytic proteins. (11) In women, estrogens elevate the risk of developing EBV-induced acquired immunodeficiency by reducing the CD4/CD8 T lymphocyte ratio, increasing the survival of B cells, promoting the release of antibodies, and increasing the expression of the major histocompatibility complex class II